
Struktur dan Ekspresi Gen
Kehidupan
ditandai oleh adanya proses metabolisme yang terjadi di dalam sel.
Metabolisme merupakan proses perubahan
kimiawi dari satu bentuk ke bentuk yang lainnya, misalnya dari bentuk yang
sederhana menjadi bentuk yang lebih rumit, atau sebaliknya. Proses metabolisme
melibatkan transformasi materi dan energi.
Penampilan morfologi yang merupakan
fenotipe dari suatu organisme adalah hasil proses metabolisme yang terjadi di dalam setiap sel penyusun
organisme tersebut. Keragaman morfologi di antara individu anggota suatu
populasi sangat tergantung dari keragaman proses dan hasil metabolisme yang
terjadi pada masing-masing individu. Perbedaan warna bunga dari satu varietas
dengan varietas lain tergantung dari proses metabolisme yang terjadi di dalam
sel dari varietas yang bersangkutan.
Proses
metabolisme di dalam sel merupakan
reaksi biokimia yang dikatalisis oleh enzim tertentu, sehingga keragaman proses
dan hasil metabolisme ditentukan oleh enzim yang terlibat dalam reaksi
tersebut. Keragaman enzim (baik struktur maupun susunan asam aminonya) itu
sendiri sangat ditentukan oleh susunan cetakannya yaitu asam deoksiribonukleat
(DNA). Ruas DNA yang menjadi cetakan untuk mensintesis enzim (protein) disebut
dengan gen, sehingga gen merupakan pengendali proses metabolisme atau
pengendali kehidupan. Keragaman morfologi suatu organisme merupakan penampakan
keragaman gen-gennya. Ilmu yang mempelajari struktur, fungsi, dan perilaku gen
disebut dengan genetika.
Struktur
gen yang dipelajari di dalam genetika meliputi struktur kimia gen, proses
pembentukannya dan pewarisannya serta perubahannya atau mutasi. Fungsi gen
dipelajari melalui peranannya di dalam sintesis protein/enzim. Pada akhirnya
genetika digunakan sebagai landasan yang bermanfaat bagi kehidupan manusia
seperti dalam pertanian, kesehatan, dan industri.
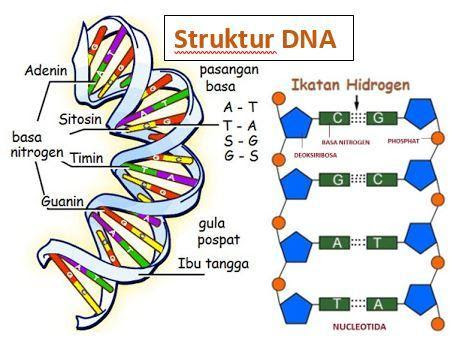
Struktur DNA
DNA merupakan bahan genetik pada semua organisme kecuali beberapa virus
tertentu yang mempunyai RNA sebagai bahan genetiknya. Bersamaan dengan
pembuktian bahwa DNA merupakan bahan genetik, penelitian tentang struktur asam
nukleat sangat intensif dilakukan karena
diharapkan pengetahuan ini dapat menjelaskan hubungan antara struktur dan
fungsi dari bahan genetik seperti dapat melakukan replikasi, merupakan
penyimpan informasi, dapat diekspresikan, dan dapat mengalami perubahan
(mutasi). Antara tahun 1940 dan 1953
banyak ahli yang tertarik untuk mengetahui struktur DNA termasuk di dalamnya adalah: Erwin Chargaff, Maurice
Wilkins, Rosalind Franklin, Linus Pauling, Francis Crick, dan James Watson. Di
antara usaha pencarian struktur DNA ini, pada tahun 1953 Watson dan Crick
mengemukakan model struktur molekul DNA yang sampai saat ini diakui
kebenarannya.
Nukleotida
Asam
nukleat merupakan molekul polimer, polinukleotida, yang tersusun dari monomer
nukleotida. Dengan demikian maka nukleotida merupakan unit dasar penyusun asam
nukleat. Nukleotida tersusun dari tiga
komponen yang penting, yaitu: basa nitrogen, gula pentosa yang
mempunyai lima karbon, dan gugus fosfat.
Ada dua jenis basa nitrogen penyusun nukleotida yaitu: basa purin yang
berstruktur cincin ganda, dan basa pirimidin yang berstruktur cincin tunggal.
Basa purin yang terdapat di dalam nukleotida ada dua tipe yaitu: adenin (A) dan
guanin (G), sedangkan 3 tipe basa pirimidin biasa terdapat di nukleotida
adalah: sitosin (C), timin (T), dan urasil (U).
DNA
dan RNA mengandung basa nitrogen A, G, C.
Basa T hanya terdapat di DNA, sedangkan basa U hanya terdapat di molekul
RNA. Gula pentosa yang terdapat di dalam
molekul RNA adalah ribosa, sedangkan
yang terdapat di molekul DNA adalah deoksiribosa.
Deoksiribosa
mirip dengan ribosa, hanya saja pada atom C2' pada deoksiribosa kekurangan gugus
hidroksil dibandingkan dengan ribosa.
Basa purin atau pirimidin yang bergabung dengan gula pentosa (ribosa
atau deoksiribosa) membentuk molekul nukleosida. Jika gugus fosfat bergabung
dengan nukleosida, maka membentuk nukleotida. Berdasarkan gula pentosanya,
terdapat dua macam nukleosida yaitu: ribonukleosida yang mengandung gula
ribosa, dan deoksiribonukleosida yang mengandung gula deoksiribosa. Nukleosida
dan nukleotida dinamai berdasarkan pada basa nitrogen yang spesifik. Ikatan
di antara tiga komponen penyusun
nukleotida adalah sangat spesifik. C1' dari gula pentosa berikatan dengan basa
nitrogen. Bila basanya purin, maka C1'
pentosa berikatan dengan N9 purin, dan bila basanya pirimidin, C1' berikatan
dengan N1 pirimidin. Pada molekul nukleotida, gugus fosfat dapat diikat pada
C2', C3', C5' dari pentosa. Bila satu gugus fosfat diikatkan pada nukleosida
akan membentuk nukleosida monofosfat, bila dua fosfat, menjadi nukleosida
difosfat, dan bila tiga fosfat menjadi nukleosida trifosfat. Bila nukleotida
tersusun dari gula ribosa, basa adenin dan tiga gugus fosfat maka disebut
dengan adenosin trifosfat (ATP). Sedangkan nukleotida yang tersusun oleh gula
deoksiribosa, basa adenin dan tiga gugus fosfat disebut dengan deoksiadenosin
trifosfat (dATP). Pada molekul nukleotida di sini, gugus fosfat diikat pada C5'
dari pentosa.
Polinukleotida
Nukleotida
yang satu diikatkan dengan nukleotida yang lain membentuk rantai polinukleotida
dengan ikatan kovalen fosfodiester. Ikatan ini terbentuk antara gugus hidroksil
(OH) pada C3' dari satu nukleotida dengan gugus fosfat pada C5' dari nukleotida
lainnya, sehingga membentuk ikatan fosfodiester 3'-5'. Nukleotida yang baru
dapat ditambahkan pada gugus OH dari C3'pada rantai nukleotida yang sedang
tumbuh.
Jadi,
penambahan nukleotida pada rantai polinukleotida selalu terletak pada gula yang
melibatkan gugus fosfat, sedangkan basa nitrogennya bebas, sehingga bentuk
polinukleotida merupakan rantai yang tersusun dari tulang punggung yang berupa
gula-fosfat dengan nitrogen sebagai percabangannya. Menurut Watson dan Crick,
DNA mempunyai utas ganda, dan kedua utas tersebut saling berpilin.
Masing-masing utas tersusun dari rantai polinukleotida. Antara polinukleotida yang
satu dengan polinukleotida yang lainnya diikat dengan ikatan-ikatan hidrogen.
Ikatan hidrogen, yang merupakan ikatan
yang lemah, terbentuk antara dua basa nitrogen. Basa guanin (G) selalu
berikatan dengan basa sitosin (C) dan basa adenin (A) selalu berikatan dengan
timin (T). Perpasangan ini didasarkan pada penelitian Chargaff, sehingga biasa
disebut dengan aturan chargaff. Chargaff telah mengisolasi DNA dari beberapa organisme, dan mendapatkan
bahwa banyaknya basa purin selalu sama dengan banyaknya basa pirimidin,
banyaknya A selalu sama dengan T, dan jumlah G sama dengan jumlah C atau
(A+G)/(T+C)=1. Jumlah G+C tidak sama dengan A+T, atau perbandingan (G+C)/(A+T)
tidak sama dengan satu tetapi bervariasi tergantung dari organismenya. Antara basa G dan C dapat terbentuk 3 ikatan
hidrogen, dan antara A dan T dihubungkan dengan 2 ikatan hidrogen, sehingga
ikatan antara G-C lebih kuat dibandingkan dengan A-T. Oleh sebab itu DNA yang
banyak mengandung G dan C lebih sulit didenaturasi (dirusak atau diuraikan) dibandingkan
dengan DNA yang banyak mengandung A dan T. Utas ganda DNA berpilin searah dengan jarum jam ke arah kanan. Dalam satu
putaran terdapat 10 pasang nukleotida. Panjang satu putaran 34 Ao sehingga
pasangan nukleotida yang satu dengan pasangan nukleotida yang berikutnya
berjarak 3,4 Ao. DNA utas ganda berpilin berdiameter 20 Ao. Utas ganda yang
berpilin ini menciptakan suatu celah. Dalam satu putaran terdapat satu celah
lebar (major groove) dan satu celah
sempit (minor groove). Struktur DNA yang
demikian, yang umumnya dijumpai disebut dengan B-DNA. Dari hasil penelitian
selanjutnya ternyata ditemukan adanya modifikasi dari struktur B-DNA.
A-DNA
mempunyai arah pilinan sama dengan B-DNA tetapi lebih kompak, yang berdiameter
23 Ao, dan dalam satu putaran terdapat 11 pasang nukleotida. Struktur lainnya
adalah Z-DNA yang mempunyai arah pilinan ke kiri (berlawanan dengan arah jarum
jam), yang mempunyai diameter 18 Ao dan setiap putaran terdapat 12 pasang
nukleotida.
Struktur
kromosom
Penyusunan DNA di dalam organisme prokaryot
berbeda dengan penyusunannya di dalam organisme eukaryot. Di dalam
organisme prokaryot atau di dalam
mitokondria dan kloroplast, DNA tidak berasosiasi dengan molekul lain. Di dalam
inti sel pada organisme eukaryot, DNA berasosiasi dengan protein histon. DNA yang berasosiasi dengan protein histon disebut
kromatin. Histon merupakan suatu protein yang bermuatan positif karena banyak
mengandung asam amino lisin dan arginin. Kromatin tersusun dari nukleosom. Satu
nukleosom tersusun dari 2 jenis molekul tetramer dan 200 pasang nukleotida serta
satu histon H1. Molekul tetramer terdiri dari 2 protein histon H2A dan 2
protein histon H2B, sedangkan tetramer lainnya tersusun dari 2 molekul protein
histon H3 dan 2 protein histon H4. Kedua tetramer ini membentuk oktomer.
Molekul oktomer ini dililit oleh DNA utas ganda sepanjang 146 pb, membentuk unsur
atau partikel inti nukleosom. Histon H1 menempel pada bagian dasar partikel
inti dan mengikat masing-masing 10 pb pada bagian kiri dan kanan dari partikel inti nukleosom.
Selain itu, nukleosom mengandung 34 pb DNA yang merupakan DNA penghubung
(linker DNA) dengan nukleosom lainnya. Jadi, dalam satu nukleosom terdapat 200
pb DNA, dan 9 molekul protein histon (1 H1, 2 H2A, 2 H2B, 2 H3, 2 H4).
Nukleosom ini mempunyai diameter 100 Ao. Nukleosom-nukleosom ini membentuk
suatu gulungan sehingga membentuk struktur selenoid atau kumparan. Setiap
gulungan terdiri dari 6 nukleosom, dan berdiameter 300 Ao.

Gulungan-gulungan ini membentuk simpul, dan
setiap 50 gulungan membentuk satu simpul yang diikat oleh suatu matriks.
Simpul-simpul ini menggulung, dan satu putaran mengandung 18 simpul yang
berdiameter 0,84 m. Simpul-simpul yang menggulung ini membentuk kromosom. Salah
satu fungsi bahan genetik adalah sebagai penyimpan informasi genetik. Sesuai
dengan fungsi ini, maka informasi yang dikandung oleh DNA harus dapat
dipertahankan atau diwariskan ke generasi berikutnya. Pewarisan informasi yang
dikandung oleh DNA dilakukan melalui proses replikasi. Replikasi atau
penggandaan DNA merupakan proses pembentukan molekul DNA yang baru berdasarkan
molekul DNA yang sudah ada. Fungsi lain dari bahan genetik adalah bahwa
informasi genetik yang dikandungnya dapat diekspresikan. Ekspresi gen dilakukan
melalui dua tahapan yaitu: transkripsi dan translasi.
Replikasi DNA
Di
dalam siklus hidupnya, sel melalui beberapa tahap yaitu: (1) tahap pertumbuhan
awal sel (G1), (2) tahap replikasi kromosom atau DNA (S), (3) tahap pertumbuhan
sel (G2), dan (4) tahap pembelahan (M). Pada tahap pertumbuhan, baik pada G1 maupun pada G2 sel mensintesis
berbagai senyawa yang dibutuhkannya seperti protein, lipid, dan karbohidrat
yang diperlukan untuk replikasi atau pembelahan sel. Pada tahap S, DNA
mengalami penggandaan dengan menggunakan berbagai senyawa yang disintesis
selama tahap G1. Setelah DNA mengalami penggandaan, sel tumbuh terus (tahap G2)
dengan mensintesis senyawa-senyawa yang diperlukan untuk tahap berikutnya,
yaitu tahap pembelahan (M). Pada tahap
G2 jumlah DNA yang terdapat di dalam sel berlipat ganda. Pada tahap M, yang
terbagi lagi ke dalam tahapan profase,
metafase, anafase, dan telofase, DNA membagi diri ke dalam dua sel anak melalui
proses pembelahan. Jadi, DNA melakukan replikasi pada saat sel akan
memperbanyak diri. DNA melakukan replikasi dengan mengikuti pola
semikonservatif.
.jpg)
Dengan
pola semikonservatif artinya bahwa setiap utas DNA menjadi cetakan bagi
pembentukan utas baru, sehingga pada akhir proses replikasi akan ditemukan dua
utas ganda yang masing-masing mengandung satu utas baru dan satu utas lama. Terdapat
dua model replikasi lainnya yaitu pola konservatif dan pola dispersif. Pada
pola konservatif, dua utas dari utas
ganda DNA secara bersama-sama membentuk dua utas baru, sehingga akan dihasilkan satu utas ganda baru
dan satu utas ganda lama. Replikasi yang mengikuti pola dispersif akan
menghasilkan dua utas ganda baru yang masing-masing terdiri dari utas lama dan
utas baru yang disusun secara selang-seling. Meselson dan Stahl telah
membuktikan bahwa kedua pola ini tidak
benar, dan yang benar adalah bahwa replikasi DNA mengikuti pola
semikonservatif. Meselson dan Stahl (1958)
melakukan percobaannya dengan menggunakan radioisotop 15N dan 14N. Unsur
nitrogen 15N berbeda dari 14N karena 15N lebih berat. Mula-mula mereka
menumbuhkan E. coli di dalam media yang mengandung 15N pada senyawa-senyawa
basa nitrogennya (generasi 0). Sebagian bakteri ini diisolasi DNAnya, sebagian
lagi ditumbuhkan pada media yang mengandung 14N. Sebagian dari generasi 1 ini
diisolasi DNAnya dan sebagian lagi dibiarkan tumbuh pada media yang sama untuk
mendapatkan generasi 2, dan seterusnya. DNA yang diisolasi dari
masing-masing generasi kemudian
disentrifugasi secara terpisah di dalam gradien kerapatan CsCl. Hasil dari
sentrifugasi menunjukkan bahwa DNA generasi 0 hanya membentuk satu pita yang
terdapat di bagian bawah tabung. Pada
generasi 1, terdapat satu pita yang letaknya berbeda dari generasi 0,
yaitu terletak sedikit di atas pita generasi 0. Ini menunjukkan bahwa DNA
generasi 1 lebih ringan dari generasi 0. Pada generasi 2, hasil sentrifugasi
menunjukkan adanya 2 pita, satu terletak pada posisi yang sama dengan generasi
1 dan satu pita lagi terdapat di
atasnya, dengan intensitas yang sama. Hal ini menunjukkan bahwa tidak terdapat
lagi utas ganda berat yang disusun hanya oleh 15N, tetapi terdapat utas-utas
ganda yang lebih ringan, yaitu yang disusun oleh 15N dan 14N atau keduanya oleh
14N.
Pada
generasi 3, hasil sentrifugasi DNA menunjukkan pola pita yang sama dengan
generasi 2 hanya saja intensitas pita yang paling atas lebih kuat dari pita
yang di tengah. Hal ini menunjukkan bahwa konsentrasi DNA utas ganda yang
tersusun dari 14N dan 14N lebih besar dari pada DNA yang tersusun dari 14N dan
15N. Pada generasi-generasi berikutnya,
pita yang di tengah makin lama makin tipis dibandingkan dengan pita yang di
atas. Hal ini menunjukkan bahwa konsentrasi utas ganda yang terdiri dari
15N/14N makin lama makin berkurang, sedangkan yang tersusun oleh 14N/14N
semakin besar konsentrasinya. Dari hasil percobaannya, Meselson dan Stahl menyimpulkan bahwa DNA melakukan
replikasi dengan pola semikonservatif. Sintesis DNA hanya bisa dimulai bila
utas ganda DNA terurai menjadi dua utas tunggal, karena utas-utas tunggal ini
yang digunakan sebagai cetakan bagi sintesis DNA yang baru. Utas ganda DNA
dapat terurai menjadi utas tunggal pada daerah yang disebut dengan titik awal
replikasi (ori). Hanya DNA yang mempunyai ori dapat melakukan replikasi. DNA
yang demikian disebut dengan replikon. Penguraian utas ganda DNA menjadi
utas-utas tunggal dilakukan oleh enzim helikase. Pada awal replikasi, helikase
menyisip pada daerah ori, kemudian diawali dari ori ini utas ganda DNA diurai menjadi dua utas tunggal.
Penguraian utas ganda ini menyebabkan tegangan pada utas ganda di atas yang
terurai. Tegangan tersebut dikurangi oleh kerja enzim DNA girase. Setelah utas
ganda DNA terurai, molekul RNA berukuran pendek disintesis dan menempel pada
ujung 3'OH dari utas DNA sesuai dengan aturan chargaff (A-T, G-C) dengan
bantuan enzim RNA polimerase atau disebut juga primase. RNA berukuran pendek
ini disebut dengan primer. Dari RNA primer ini DNA disintesis dengan menggunakan cetakan utas DNA yang sudah ada
dengan arah sintesis dari 5'P ke 3'OH dan mengikuti aturan chargaff. Sintesis
DNA yang berawal dari RNA primer dikalisis oleh enzim DNA polimerase III. DNA utas ganda yang diuraikan helikase
menimbulkan bentuk garpu, dimana percabangan garpu berakhir pada DNA yang belum
terurai.
Utas
tunggal hasil penguraian utas ganda oleh
enzim helikase dipertahankan kestabilannya oleh protein pengikat utas tunggal SSBP (single-stranded binding
protein). Sintesis DNA mempunyai arah 5' ke 3' sehingga pada saat percabangan
garpu memanjang, maka pada salah satu utas akan disintesis DNA secara kontinyu,
sedangkan pada utas lainnya sintesis DNA tidak kontinyu. Utas DNA yang
digunakan sebagai cetakan bagi DNA yang
disintesis secara kontinyu disebut dengan utas leading, sedangkan utas DNA yang
digunakan sebagai cetakan bagi DNA yang disintesis secara tidak kontinyu
disebut dengan utas lagging. Pada utas lagging terdapat beberapa fragmen atau potongan
DNA. Potongan-potongan ini disebut
dengan fragmen Okazaki. Antara satu
fragmen dan fragmen lainnya terdapat celah. DNA polimerase I berfungsi untuk
menghilangkan RNA primer dan sekaligus mensintesis DNA pada celah di antara dua
fragmen Okazaki. Setelah disintesis oleh DNA polimerase I, kedua potongan
DNA yang sudah berdekatan disambungkan
satu dengan lainnya oleh enzim ligase.
Ekspresi Gen
Ekspresi gen adalah proses penentuan sifat
dari suatu organisme oleh gen. Suatu sifat yang dipunyai oleh suatu organisme
merupakan hasil proses metabolisme yang terjadi di dalam sel. Proses
metabolisme dapat berlangsung karena adanya enzim yang berfungsi sebagai
katalisator proses-proses biokimia. Enzim dan protein lainnya diterjemahkan
dari urutan nukleotida yang ada pada molekul mRNA, dan mRNA itu sendiri disintesis berdasarkan utas
cetakan DNA. Gen tersusun dari molekul
DNA, sehingga gen menentukan sifat suatu organisme. Seperti telah disinggung di depan bahwa
ekspresi gen dilakukan memalui dua tahap, yaitu: transkripsi dan translasi.
Proses transkripsi terjadi di dalam inti sel, sedangkan translasi berlangsung
di sitoplasma, sehingga RNA harus dikeluarkan dari inti sel ke sitoplasma.
Transkripsi
Transkripsi
merupakan proses pembentukan molekul RNA
dengan menggunakan DNA sebagai
cetakannya. Tidak semua bagian DNA akan ditranskripsikan, tetapi hanya
bagian tertentu saja. Bagian tertentu tersebut disebut dengan gen.
Keseluruhan DNA baik gen maupun sekuensi
DNA bukan penyandi (non-coding) yang dikandung oleh suatu organisme disebut
dengan genom. Ruas DNA yang akan
ditranskripsikan dibatasi oleh promoter dan terminator. Hanya satu dari dua
utas DNA yang digunakan sebagai cetakan sintesis RNA. Utas DNA yang digunakan
sebagai cetakan bagi sintesis RNA disebut dengan utas cetakan (template),
sedangkan utas DNA lainnya disebut dengan utas pendamping. Walaupun hanya satu
utas yang berfungsi sebagai cetakan, tetapi tidak selalu utas yang sama digunakan sebagai utas cetakan sepanjang
molekul DNA di dalam genom suatu
organisme. Jadi, pada satu gen, utas yang satu digunakan sebagai cetakan,
tetapi pada gen lainnya, kemungkinan utas yang lain digunakan sebagai cetakan.
Proses transkripsi menghasilkan tiga jenis RNA, yaitu: RNA duta
(mRNA= messenger RNA), RNA transfer (tRNA= transfer RNA), dan RNA ribosomal
(rRNA= ribosomal RNA). Ketiga jenis RNA ini berperanan di dalam proses
translasi. Hanya mRNA yang akan diterjemahkan kedalam protein. tRNA berperan
sebagai molekul pembawa asam amino yang akan dirangkaikan menjadi polipeptida
sesuai dengan sandi yang terdapat pada mRNA. rRNA berfungsi sebagai salah satu
molekul penyusun ribosom. Proses transkripsi dikatalisis oleh enzim
transkriptase atau RNA polimerase. Pada organisme prokaryot seperti E. coli,
hanya terdapat satu jenis RNA polimerase untuk mengkatalisis sintesis semua
jenis RNA. Pada organisme eukaryot, terdapat tiga jenis RNA polimerase, yaitu:
(1) RNA polimerase I yang berfungsi untuk mengkatalisis pembentukan rRNA, (2)
RNA polimerase II yang berperan dalam sintesis tRNA dan beberapa molekul rRNA,
dan (3) RNA polimerase III yang bertugas mengkalisis proses sintesis mRNA.
Enzim RNA polimerase lengkap (disebut holoenzim) tersusun dari enzim inti dan
faktor transkripsi.
Enzim
inti tersusun dari dua subunit. Proses transkripsi mempunyai beberapa
karakteristik yaitu bahwa: proses sintesis mempunyai arah dari 5'P ke 3'OH,
berlangsung secara anti paralel bila dibandingkan dengan utas cetakannya, dan mengikuti aturan chargaff atau
basa-basanya berpasangan secara
komplementer (A-T; G-C). Proses transkripsi dapat dibagi kedalam tiga
tahap, yaitu: inisiasi sintesis RNA, pemanjangan (elongasi) RNA dan
penyelesaian (terminasi) sinteis RNA. Inisiasi dimulai pada saat RNA polimerase
menempel pada molekul DNA utas ganda. RNA polimerase dapat menempel pada situs
DNA secara spesifik karena adanya faktor sigma. Pada daerah penempelan RNA
polimerase ini, utas ganda DNA terurai menjadi utas tunggal secara lokal.
Nukleotida pertama kemudian ditempatkan di depan utas cetakan kemudian
disintesis nukleotida berikutnya sesuai dengan utas cetakan yang ada
didepannya. Titik atau situs dimana nukleotida pertama diletakkan di depan utas
cetakan disebut dengan titik atau situs inisiasi. Setelah terbentuk rangkaian
RNA yang tersusun dari 2-9 nukleotida, faktor sigma meninggalkan kompleks enzim
inti. Promoter adalah situs (daerah) yang dikenali pertama kali oleh RNA
polimerase sebagai tempat penempelannya. Promoter ini merupakan sekuensi DNA
yang terdiri dari sekitar 40 pb yang terletak tepat sebelum situs mulainya
transkripsi. Untuk memudahkan pada ahli, pemberian kode +1 berarti bahwa pada
situs tersebut nukleotida pertama
disintesis. Jadi, promoter terletak diaerah sebelum atau di depan
(up-stream) situs +1. Daerah sebelum +1 diberi kode mulai -1, -2, dan
seterusnya. Pada prokaryot, dua daerah yang hampir selalu ada pada promoter
adalah sekuensi -35 dan -10. Kedua daerah tersebut terdiri dari 6 pb yang
keduanya dipisahkan oleh sekitar 25 pb. Pada daerah -35 (antara -35 - -30),
tiga basa yang hampir selalu (75%) ada
adalah: TTG.... Pada daerah -10 (antara
-12, - 7), basa yang dikonservasi adalah: TA...T. Daerah ini disebut juga
dengan kotak Pribnow (Pribnow box) yang merupakan daerah peringatan bagi RNA
polimerase untuk memulai transkripsi.
Pada
eukaryot, RNA polimerase II akan
mengenali daerah promoter yang lebih awal yaitu pada -130 yang disebut juga
dengan kotak CAAT (CAAT box) dan -30 yang disebut dengan kotak TATA (TATA box)
atau disebut juga kotak Goldberg-Hogness yang kaya basa AT. Kotak TATA ini
mempunyai kesamaan dengan kotak Pribnow pada prokaryot. Proses pemanjangan RNA
dilakukan oleh RNA ploimerase yang sudah tidak mengandung faktor sigma. Pada
tahap ini ribonukleotida secara suksesif menempel pada utas RNA yang sedang tumbuh
membentuk hibrid DNA/RNA. RNA polimerase
bergerak terus sepanjang utas DNA sambil memisahkan kedua utas DNA. Utas DNA
yang terurai secara lokal ini besarnya
sekitar 17 pb. Pada tahap pemanjangan ini RNA polimerase menutupi DNA sepanjang sekitar 60 pb. Dengan bergeraknya
sintesis RNA, pada bagian tertentu dari utas RNA yang telah disintesis berpisah
dengan utas DNA, sedangkan bagian lain masih membentuk hibrid RNA/DNA dengan
panjang sekitar 12 pb. Pada daerah terminator, RNA polimerase tidak mampu lagi
menempelkan ribonukleotida pada RNA yang telah terbentuk. Pada saat ini disebut
dengan tahap terminasi. Pada tahap ini NusA diganti oleh faktor rho. Pada tahap
ini RNA polimerase dan RNA dibebaskan dari DNA, dan DNA membentuk utas ganda
kembali. Terminator pada prokaryot ditandai oleh daerah yang simetri tidak
sempurna, dan daerah tersebut biasanya kaya basa AT yang mengikuti daerah
simetri tidak sempurna.
Translasi
Dalam proses translasi asam amino akan
dirangkaikan dengan asam amino lainnya untuk membentuk rantai polipeptida atau
protein. Jenis asam amino amino yang dirangkaikan ditentukan oleh urutan
nukleotida yang terdapat pada molekul mRNA. Jadi, mRNA digunakan sebagai model
cetakan bagi sintesis protein. Asam amino
dirangkaikan dengan asam amino
lain dengan ikatan peptida yang dilakukan oleh ribosom. Asam amino yang akan dirangkaikan dengan asam
amino lainnya dibawa oleh tRNA. Setiap asam amino akan dibawa oleh tRNA yang
spesifik ke dalam kompleks mRNA-ribosom.
mRNA merupakan
rangkaian kodon yang akan dibaca oleh ribosom. Kodon pada mRNA akan
berpasangan dengan antikodon yang ada pada tRNA. Setiap tRNA mempunyai antikodon
yang spesifik. Translasi berlangsung mulai dari kodon awal sampai kodon akhir. Hubungan antara kodon
dengan asam amino diatur melalui sandi genetik. Dalam proses translasi ini
hanya ada satu kodon awal yaitu AUG yang
menyandi asam amino metionin dan tiga kodon akhir: UAA, UAG, dan UGA. Seperti
pada proses transkripsi, proses translasi dapat dibagi ke dalam tiga tahap: inisiasi,
pemanjangan, dan penyelesaian. Pada tahap inisiasi, ribosom akan menempel pada
mRNA pada daerah yang spesifik. Ribosom mempunyai dua situs penempelan untuk
tRNA, yaitu situs P (peptidil) dan situs A (aminoasil). Bilamana ribosom ini
bertemu dengan kodon awal (AUG pada mRNA), maka tRNA yang membawa metionin akan
masuk ke dalam situs P di dalam ribosom, dan ribosom akan membaca kodon
disebelahnya (yang ada di bawahnya).
Sesuai dengan kodonnya, tRNA yang membawa asam amino tertentu akan
memasuki situs A. Proses pemanjangan dimulai bilamana ribosom bergerak ke bawah
(ke arah 3'OH). tRNA yang tadinya berada pada situs P akan keluar dari kompleks
ribosom-mRNA sambil memindahkan asam amino yang dibawanya kepada tRNA yang
berada pada situs P yang tadinya berada pada situs A. Pada saat yang bersamaan situs A menjadi
kosong. Situs yang kosong ini akan diisi oleh tRNA yang membawa asam amino
tertentu. Bilamana ribosom ini bergerak lagi ke bawah sambil membaca kodon berikutnya,
tRNA yang berada pada situs P keluar dari situs tersebut sambil memindahkan
polipeptida yang sedang tumbuh yang dibawanya ke pada asam amino yang dibawa
oleh tRNA yang berada pada situs P yang
berasal dari situs A. Situs A akan diisi oleh tRNA yang baru lagi. Ribosom ini
akan bergerak terus dengan arah 5'P ke 3'OH sepanjang mRNA sambil merangkaian
asam amino. Proses penyelesaian atau terminasi ditandai bila ribosom bertemu
dengan kodon akhir. Pada saat ini tidak satupun asam amino yang dirangkaikan
sehingga proses sintesis protein berakhir.
Ribosom
kemudian berpisah dari mRNA dan terurai menjadi 2 subunit, yaitu sub unit besar
dan sub unit kecil. Selama proses translasi, subunit kecil menempel pada mRNA
sedangkan subunit besar berperan sebagai tempat tRNA (situs P dan situs
A). Contoh proses pembentukan protein
dari molekul DNA secara sederhana:
U.
pendamping : 5'-A T G G G T A C C C A T
G C T T T T G C C-3'
U.
cetakan : 3'-T A C C C A T G G
G T A C G A A A A C G G-5'
mRNA : 5'-A U G G G U A C C C A U G C U U U U
G C C-3'
Protein : Met - Gly - Thr - His - Ser - Phe - Ala -

.jpg)


.jpg)

No comments