
RANGKAI KELAMIN DAN PENENTUAN JENIS KELAMIN
Keberadaan gen berangkai pada suatu spesies organisme, yang meliputi urutan dan jaraknya satu sama lain, menghasilkan peta kromosom untuk spesies tersebut, misalnya peta kromosom pada lalat Drosophila melanogaster yang terdiri atas empat kelompok gen berangkai.
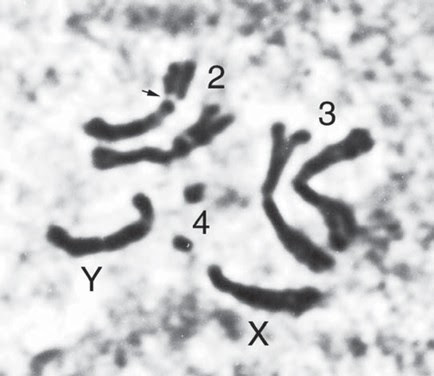
Salah satu dari keempat kelompok gen berangkai atau keempat pasang kromosom pada D. melanogaster tersebut, dalam hal ini kromosom nomor 1, disebut sebagai kromosom kelamin. Pemberian nama ini karena strukturnya pada individu jantan dan individu betina memperlihatkan perbedaan sehingga dapat digunakan untuk membedakan jenis kelamin individu. Ternyata banyak sekali spesies organisme lainnya, terutama hewan dan juga manusia, mempunyai kromosom kelamin.
Gen-gen yang terletak pada kromosom kelamin dinamakan gen rangkai kelamin (sex-linked genes) sementara fenomena yang melibatkan pewarisan gen-gen ini disebut peristiwa rangkai kelamin (linkage). Adapun gen berangkai yang dibicarakan pada Bab V adalah gen-gen yang terletak pada kromosom selain kromosom kelamin, yaitu kromosom yang pada individu jantan dan betina sama strukturnya sehingga tidak dapat digunakan untuk membedakan jenis kelamin. Kromosom semacam ini dinamakan autosom.
Seperti halnya gen berangkai (autosomal), gen-gen rangkai kelamin tidak mengalami segregasi dan penggabungan secara acak di dalam gamet-gamet yang terbentuk. Akibatnya, individu-individu yang dihasilkan melalui kombinasi gamet tersebut memperlihatkan nisbah fenotipe dan genotipe yang menyimpang dari hukum Mendel. Selain itu, jika pada percobaan Mendel perkawinan resiprok (genotipe tetua jantan dan betina dipertukarkan) menghasilkan keturunan yang sama, tidak demikian halnya untuk sifat-sifat yang diatur oleh gen rangkai kelamin.
Gen rangkai kelamin dapat dikelompok-kelompokkan berdasarkan atas macam kromosom kelamin tempatnya berada. Oleh karena kromosom kelamin pada umumnya dapat dibedakan menjadi kromosom X dan Y, maka gen rangkai kelamin dapat menjadi gen rangkai X (X-linked genes) dan gen rangkai Y (Y-linked genes). Di samping itu, ada pula beberapa gen yang terletak pada kromosom X tetapi memiliki pasangan pada kromosom Y. Gen semacam ini dinamakan gen rangkai kelamin tak sempurna (incompletely sex-linked genes). Pada bab ini akan dijelaskan cara pewarisan macam-macam gen rangkai kelamin tersebut serta beberapa sistem penentuan jenis kelamin pada berbagai spesies organisme.
Pewarisan Rangkai X
Percobaan yang pertama kali mengungkapkan adanya peristiwa rangkai kelamin dilakukan oleh T.H Morgan pada tahun 1910. Dia menyilangkan lalat D. melanogaster jantan bermata putih dengan betina bermata merah. Lalat bermata merah lazim dianggap sebagai lalat normal atau tipe alami (wild type), sedang gen pengatur tipe alami, misalnya pengatur warna mata merah ini, dapat dilambangkan dengan tanda +. Biasanya, meskipun tidak selalu, gen tipe alami bersifat dominan terhadap alel mutannya.
Hasil persilangan Morgan tersebut, khususnya pada generasi F1, ternyata berbeda jika tetua jantan yang digunakan adalah tipe alami (bermata merah) dan tetua betinanya bermata putih. Dengan perkataan lain, perkawinan resiprok menghasilkan keturunan yang berbeda. Persilangan resiprok dengan hasil yang berbeda ini memberikan petunjuk bahwa pewarisan warna mata pada Drosophila ada hubungannya dengan jenis kelamin, dan ternyata kemudian memang diketahui bahwa gen yang mengatur warna mata pada Drosophila terletak pada kromosom kelamin, dalam hal ini kromosom X. Oleh karena itu, gen pengatur warna mata ini dikatakan sebagai gen rangkai X. akan nampak bahwa lalat F1 betina mempunyai mata seperti tetua jantannya, yaitu normal/merah. Sebaliknya, lalat F1 jantan warna matanya seperti tetua betinanya, yaitu putih. Pewarisan sifat semacam ini disebut sebagai criss cross inheritance.
Pada Drosophila, dan juga beberapa spesies organisme lainnya, individu betina membawa dua buah kromosom X, yang dengan sendirinya homolog, sehingga gamet-gamet yang dihasilkannya akan mempunyai susunan gen yang sama. Oleh karena itu, individu betina ini dikatakan bersifat homogametik. Sebaliknya, individu jantan yang hanya membawa sebuah kromosom X akan menghasilkan dua macam gamet yang berbeda, yaitu gamet yang membawa kromosom X dan gamet yang membawa kromosom Y. Individu jantan ini dikatakan bersifat heterogametik.
Rangkai X pada kucing
Warna bulu pada kucing ditentukan oleh suatu gen rangkai X. Dalam keadaan heterozigot gen ini menyebabkan warna bulu yang dikenal dengan istilah tortoise shell. Oleh karena genotipe heterozigot untuk gen rangkai X hanya dapat dijumpai pada individu betina, maka kucing berbulu tortoise shell hanya terdapat pada jenis kelamin betina. Sementara itu, individu homozigot dominan (betina) dan hemizigot dominan (jantan) mempunyai bulu berwarna hitam. Individu homozigot resesif (betina) dan hemizigot resesif (jantan) akan berbulu kuning.
Istilah hemizigot digunakan untuk menyebutkan genotipe individu dengan sebuah kromosom X. Individu dengan gen dominan yang terdapat pada satu-satunya kromosom X dikatakan hemizigot dominan. Sebaliknya, jika gen tersebut resesif, individu yang memilikinya disebut hemizigot resesif.
Rangkai X pada manusia
Salah satu contoh gen rangkai X pada manusia adalah gen resesif yang menyebabkan penyakit hemofilia, yaitu gangguan dalam proses pembekuan darah. Sebenarnya, kasus hemofilia telah dijumpai sejak lama di negara-negara Arab ketika beberapa anak laki-laki meninggal akibat perdarahan hebat setelah dikhitan. Namun, waktu itu kematian akibat perdarahan ini hanya dianggap sebagai takdir semata.
Hemofilia baru menjadi terkenal dan dipelajari pola pewarisannya setelah beberapa anggota keluarga Kerajaan Inggris mengalaminya. Awalnya, salah seorang di antara putra Ratu Victoria menderita hemofilia sementara dua di antara putrinya karier atau heterozigot. Dari kedua putri yang heterozigot ini lahir tiga cucu laki-laki yang menderita hemofilia dan empat cucu wanita yang heterozigot. Melalui dua dari keempat cucu yang heterozigot inilah penyakit hemofilia tersebar di kalangan keluarga Kerajaan Rusia dan Spanyol. Sementara itu, anggota keluarga Kerajaan Inggris saat ini yang merupakan keturunan putra/putri normal Ratu Victoria bebas dari penyakit hemofilia.
Rangkai Z pada ayam
Pada dasarnya pola pewarisan sifat rangkai Z sama dengan pewarisan sifat rangkai X. Hanya saja, kalau pada rangkai X individu homogametik berjenis kelamin pria/jantan sementara individu heterogametik berjenis kelamin wanita/betina, pada rangkai Z justru terjadi sebaliknya. Individu homogametik (ZZ) adalah jantan, sedang individu heterogametik (ZW) adalah betina.
Contoh gen rangkai Z yang lazim dikemukakan adalah gen resesif br yang menyebabkan pemerataan pigmentasi bulu secara normal pada ayam. Alelnya, Br, menyebabkan bulu ayam menjadi burik. Jadi, pada kasus ini alel resesif justru dianggap sebagai tipe alami atau normal (dilambangkan dengan +), sedang alel dominannya merupakan alel mutan.
Pewarisan Rangkai Y
Pada umumnya kromosom Y hanya sedikit sekali mengandung gen yang aktif. Jumlah yang sangat sedikit ini mungkin disebabkan oleh sulitnya menemukan alel mutan bagi gen rangkai Y yang dapat menghasilkan fenotipe abnormal. Biasanya suatu gen/alel dapat dideteksi keberadaannya apabila fenotipe yang dihasilkannya adalah abnormal. Oleh karena fenotipe abnormal yang disebabkan oleh gen rangkai Y jumlahnya sangat sedikit, maka gen rangkai Y diduga merupakan gen yang sangat stabil.
Gen rangkai Y jelas tidak mungkin diekspresikan pada individu betina/wanita sehingga gen ini disebut juga gen holandrik. Contoh gen holandrik pada manusia adalah Hg dengan alelnya hg yang menyebabkan bulu kasar dan panjang, Ht dengan alelnya ht yang menyebabkan pertumbuhan bulu panjang di sekitar telinga, dan Wt dengan alelnya wt yang menyebabkan abnormalitas kulit pada jari.
Pewarisan Rangkai Kelamin Tak Sempurna
Meskipun dari uraian di atas secara tersirat dapat ditafsirkan bahwa kromosom X tidak homolog dengan kromosom Y, ternyata ada bagian atau segmen tertentu pada kedua kromosom tersebut yang homolog satu sama lain. Dengan perkataan lain, ada beberapa gen pada kromosom X yang mempunyai alel pada kromosom Y. Pewarisan sifat yang diatur oleh gen semacam ini dapat dikatakan tidak dipengaruhi oleh jenis kelamin, dan berlangsung seperti halnya pewarisan gen autosomal. Oleh karena itu, gen-gen pada segmen kromosom X dan Y yang homolog ini disebut juga gen rangkai kelamin tak sempurna.
Pada D. melanogaster terdapat gen rangkai kelamin tak sempurna yang menyebabkan pertumbuhan bulu pendek.
Dapat dilihat pada Gambar 6.2 bahwa perkawinan resiprok untuk gen rangkai kelamin tak sempurna akan memberikan hasil yang sama seperti halnya hasil yang diperoleh dari perkawinan resiprok untuk gen-gen autosomal. Jadi, pewarisan gen rangkai kelamin tak sempurna mempunyai pola seperti pewarisan gen autosomal.
Sistem Penentuan Jenis Kelamin
Telah disebutkan di atas bahwa pada manusia dan mamalia, dalam hal ini kucing, individu pria/jantan adalah heterogametik (XY) sementara wanita/betina adalah homogametik (XX). Sebaliknya, pada ayam individu jantan justru homogametik (ZZ) sementara individu betinanya heterogametik (ZW). Penentuan jenis kelamin pada manusia/mamalia dikatakan mengikuti sistem XY, sedang pada ayam, dan unggas lainnya serta ikan tertentu, mengikuti sistem ZW.
Selain kedua sistem tersebut, masih banyak sistem penentuan jenis kelamin lainnya. Berikut ini akan dijelaskan beberapa di antaranya.
Sistem XO
Sistem XO dijumpai pada beberapa jenis serangga, misalnya belalang. Di dalam sel somatisnya, individu betina memiliki dua buah kromosom X sementara individu jantan hanya mempunyai sebuah kromosom X. Jadi, hal ini mirip dengan sistem XY. Bedanya, pada sistem XO individu jantan tidak mempunyai kromosom Y. Dengan demikian, jumlah kromosom sel somatis individu betina lebih banyak daripada jumlah pada individu jantan. Sebagai contoh, E.B. Wilson menemukan bahwa sel somatis serangga Protenor betina mempunyai 14 kromosom, sedang pada individu jantannya hanya ada 13 kromosom.
Sistem nisbah X/A
C.B. Bridge melakukan serangkaian penelitian mengenai jenis kelamin pada lalat Drosophila. Dia berhasil menyimpulkan bahwa sistem penentuan jenis kelamin pada organisme tersebut berkaitan dengan nisbah banyaknya kromosom X terhadap banyaknya autosom, dan tidak ada hubungannya dengan kromosom Y. Dalam hal ini kromosom Y hanya berperan mengatur fertilitas jantan. Secara ringkas penentuan jenis kelamin dengan sistem X/A pada lalat Drosophila dapat dilihat pada Tabel 6.1.
Tabel 6.1. Penentuan jenis kelamin pada lalat Drosophila
| Σ kromosom X | Σ autosom | nibah X/A | jenis kelamin |
| 1 | 2 | 0,5 | jantan |
| 2 | 2 | 1 | betina |
| 3 | 2 | 1,5 | metabetina |
| 4 | 3 | 1,33 | metabetina |
| 4 | 4 | 1 | betina 4n |
| 3 | 3 | 1 | betina 3n |
| 3 | 4 | 0,75 | interseks |
| 2 | 3 | 0,67 | interseks |
| 2 | 4 | 0,5 | jantan |
| 1 | 3 | 0,33 | metajantan |
Jika kita perhatikan kolom pertama pada Tabel 6.1 akan terlihat bahwa ada beberapa individu yang jumlah kromosom X-nya lebih dari dua buah, yakni individu dengan jenis kelamin metabetina, betina triploid dan tetraploid, serta interseks. Adanya kromosom X yang didapatkan melebihi jumlah kromosom X pada individu normal (diploid) ini disebabkan oleh terjadinya peristiwa yang dinamakan gagal pisah (non disjunction), yaitu gagal berpisahnya kedua kromosom X pada waktu pembelahan meiosis.
Pada Drosophila terjadinya gagal pisah dapat menyebabkan terbentuknya beberapa individu abnormal seperti nampak pada Gambar 6.3.
gagal pisah
gamet : AXX AO AX AY
F1 : AAXXX AAXXY AAXO AAOY
betina super betina jantan steril letal
Gambar 6.3. Diagram munculnya beberapa individu abnormal pada
Drosophila akibat peristiwa gagal pisah
Di samping kelainan-kelainan tersebut pernah pula dilaporkan adanya lalat Drosophila yang sebagian tubuhnya memperlihatkan sifat-sifat sebagai jenis kelamin jantan sementara sebagian lainnya betina. Lalat ini dikatakan mengalami mozaik seksual atau biasa disebut dengan istilah ginandromorfi. Penyebabnya adalah ketidakteraturan distribusi kromosom X pada masa-masa awal pembelahan mitosis zigot. Dalam hal ini ada sel yang menerima dua kromosom X tetapi ada pula yang hanya menerima satu kromosom X.
Partenogenesis
Pada beberapa spesies Hymenoptera seperti semut, lebah, dan tawon, individu jantan berkembang dengan cara partenogenesis, yaitu melalui telur yang tidak dibuahi. Oleh karena itu, individu jantan ini hanya memiliki sebuah genom atau perangkat kromosomnya haploid.
Sementara itu, individu betina dan golongan pekerja, khususnya pada lebah, berkembang dari telur yang dibuahi sehingga perangkat kromosomnya adalah diploid. Dengan demikian, dapat dikatakan bahwa partenogenesis merupakan sistem penentuan jenis kelamin yang tidak ada sangkut pautnya sama sekali dengan kromosom kelamin tetapi hanya bergantung kepada jumlah genom (perangkat kromosom).
Sistem gen Sk-Ts
Di atas disebutkan bahwa sistem penentuan jenis kelamin pada lebah tidak berhubungan dengan kromosom kelamin. Meskipun demikian, sistem tersebut masih ada kaitannya dengan jumlah perangkat kromosom.
Pada jagung dikenal sistem penentuan jenis kelamin yang tidak bergantung, baik kepada kromosom kelamin maupun jumlah genom, tetapi didasarkan atas keberadaan gen tertentu. Jagung normal monosius (berumah satu) mempunyai gen Sk, yang mengatur pembentukan bunga betina, dan gen Ts, yang mengatur pembentukan bunga jantan. Jagung monosius ini mempunyai fenotipe Sk_Ts_.
Sementara itu, alel-alel resesif sk dan ts masing-masing menghalangi pembentukan bunga betina dan mensterilkan bunga jantan. Oleh karena itu, jagung dengan fenotipe Sk_tsts adalah betina diosius (berumah dua), sedang jagung skskTs_ adalah jantan diosius. Jagung sksktsts berjenis kelamin betina karena ts dapat mengatasi pengaruh sk, atau dengan perkataan lain, bunga betina tetap terbentuk seakan-akan tidak ada alel sk.
Pengaruh lingkungan
Sistem penentuan jenis kelamin bahkan ada pula yang bersifat nongenetik. Hal ini misalnya dijumpai pada cacing laut Bonellia, yang jenis kelaminnya semata-mata ditentukan oleh faktor lingkungan.. F. Baltzer menemukan bahwa cacing Bonellia yang berasal dari sebuah telur yang diisolasi akan berkembang menjadi individu betina. Sebaliknya, cacing yang hidup di lingkungan betina dewasa akan mendekati dan memasuki saluran reproduksi cacing betina dewasa tersebut untuk kemudian berkembang menjadi individu jantan yang parasitik.






No comments